Modeling plankton communities in the global ocean
A key part of the Darwin Project is developing theoretical and numerical models of the marine ecosystems. We specifically develop models to help us understand the diversity and biogeography of the plankton communities using trait based approaches. These models are helping us explore the role of plankton in climate change, mixotrophy, nitrogen fixation, and top-down control.
Our numerical models have been developed within the MITgcm framework, and have also been implemented in ROMS (Goebel et al. 2013) and the NCAR CESM. The code is available with example experiments at MITgcm_contrib/darwin2. In particular there are two types of models: “Monod” and “Quota”. The former uses simple monod kinetics (e.g. Follows et al. 2007; Dutkiewicz et al. 2009), while the latter incorporates internal stores of elements in a Droop-style model (e.g. Ward et al 2012). We are however working to incorporate more physiological detail in terms of a “macro-molecular model“.
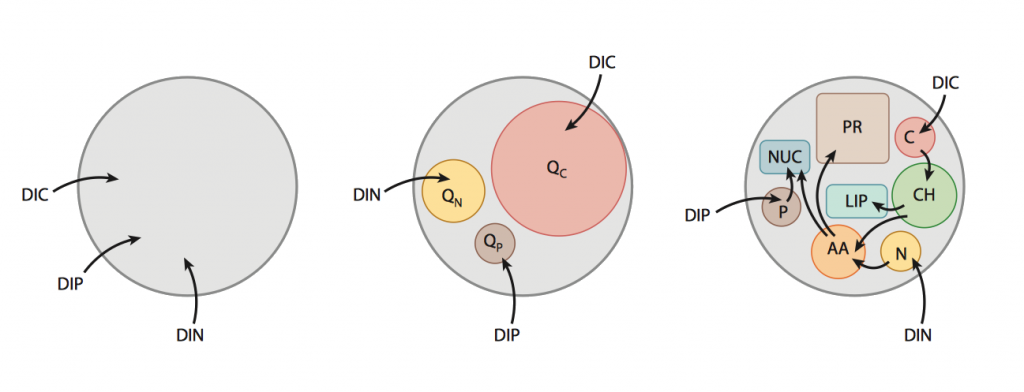
- Schematic representation of the parameterization of photoautotroph physiology employed in marine ecosystem models: (left) Monod- type, (center) Droop/Caperon-type with individual quota for each element, (right) schematic concept for a model of algal physiology that resolves key biochemical components of an algal cell. Image from Follows and Dutkiewicz, 2011
Significant developments of our models are recorded in the papers:
- Follows et al 2007
- Dutkiewicz et al 2009
- Monteiro et al 2010
- Hickman et al 2010
- Follows and Dutkiewicz 2011
- Ward et al 2012
Diversity and Biogeography
Phytoplankton are an extremely diverse set of organisms spanning nine orders of magnitude in cell size and an enormous range of morphologies, biochemical function, elemental requirements, and trophic strategy. The type of assemblage of phytoplankton has a key role in regulating biogeochemical pathways and fluxes including the export of organic matter to the deep ocean. We explore what sets the diversity of the assemblages (e.g. null hypothesis, dispersion, trait strategies, top down control) and what sets the biogeography of these assemblages. Explore →
Trait Based Approaches: size and biogeochemical function
Phytoplankton assemblages have been empirically shown to vary systematically in size, depending in part on resource delivery rates, with important ecological and biogeochemical consequences, for example modulating export. Biogeochemical functionality of assemblages also shows systematic variations with impact on biogeochemical cycles. We explore methods to model and understand phytoplankton communities from trait-based approaches.
- Cell size regulates several key characteristics such as maximum growth rate, nutrient affinity, maximal elemental cell quotas, sinking speeds and susceptibility to grazing. These relationships can be interpreted biophysically, invoking the scaling with size and encounter rates, fluxes through molecular boundary layers or internal redistribution networks.
- Functional traits reflect specific biochemical pathways and functions and are determined by how the cell invests energy and elemental resources. Some key functional traits include, for example, the ability to fix dinitrogen gas and the use of specific minerals (e.g. calcium carbonate or silicate), differences in elemental composition, different optimal temperature, and light adaptation, morphology, predator avoidance and variation of trophic strategies.
Nitrogen Fixation
Nitrogen fixers or diazotrophs, organisms that fix nitrogen gas (N2) into an organic form, play an important role in the climate system as the availability of inorganic nitrogen can represent a significant limiting factor for marine primary production. On the global scale, diazotrophy is the largest source of fixed nitrogen in the modern ocean counterbalancing the sinks due to denitrification and anammox. We have used our models and resource competition theory to understand the geographic distribution of diazotrophs in the current ocean, and potential changes in a future ocean. We have also explored the importance of a diversity of diazotroph types. Explore →
Top-Down Control
Top-down mechanisms by consumers (predators) can shape the phytoplankton community structure. Using numerical simulations and theory we explore the importance of top-down controls on the size distribution of phytoplankton as well as “kill-the winner” dynamics in controlling the diversity of phytoplankton. Explore →
Climate Change
Ocean ecosystems are increasingly impacted by human-induced changes to their environment. The oceans are warming, acidifying, and mixing and circulation are changing. It is likely that the physical changes will reduce rates of nutrient supply to the euphotic zone and select for smaller phytoplankton cells better adapted to oligotrophic conditions. Such shifts are likely to impact the export of carbon to the deep ocean. We explore the individual and combined effect of how multiple stressors act on the phytoplankton communities. We explore how incorporating the diversity of ecosystems is crucial for understanding potential changes to the phytoplankton communities, biogeochemical functioning, and stability. Explore →
Mixotrophy
Mixotrophic organisms combine autotrophic and heterotrophic nutrition. Recent observations indicate that mixotrophs constitute a large fraction of the biomass, bacterivory, and primary production in oligotrophic environments. While mixotrophy allows greater flexibility in terms of resource acquisition, any advantage must be traded off against an associated increase in metabolic costs, which appear to make mixotrophs uncompetitive relative to specialist autotrophs and heterotrophs. We use observations, models and simple theory to identify when and where mixotrophy is a useful strategy. Explore →
Macromolecular Modeling
The parameterizations of microbial physiology in current ocean ecosystem models are constructed using 1940’s modeling paradigms. We are working on a new paradigm that resolves key macro-molecular pools in the cell and employs powerful constraints from mass, redox, and energy conservation. This framework is developed to be sufficiently general to describe a wide variety of microbial lifestyles, computationally tractable for global ocean simulations, and yet scalable to employ and interface with, genome-scale information. Our prototype model currently examines the cost of trait-trade-offs, especially for nitrogen fixation organisms. Explore →